
撰文 | 刘开杰(2023级博士研究生)
审稿 | 梅林
人们很早就认识到运动对身体的影响。伏尔泰就说过,生命在于运动。研究发现,运动对于我们身体有许多好处,例如促进骨骼发育、增强心肺功能,降低糖尿病、心血管疾病以及抑郁和焦虑等疾病风险,提高意识能力等。我国西汉早期就有通过成体系的体操运动以增强体质的记录。在本世纪之前,关于运动对健康的影响主要集中在调查与相关性分析。二十一世纪以来,科学家们开始研究运动有利健康的病理分子机制,并且认为运动对个体益处可能通过细胞,尤其是肌肉细胞,分泌到血液中的一些因子而实现的。随着代谢组学的发展,科学家们在运动后人体血液中检测到了很多和代谢相关的分子。
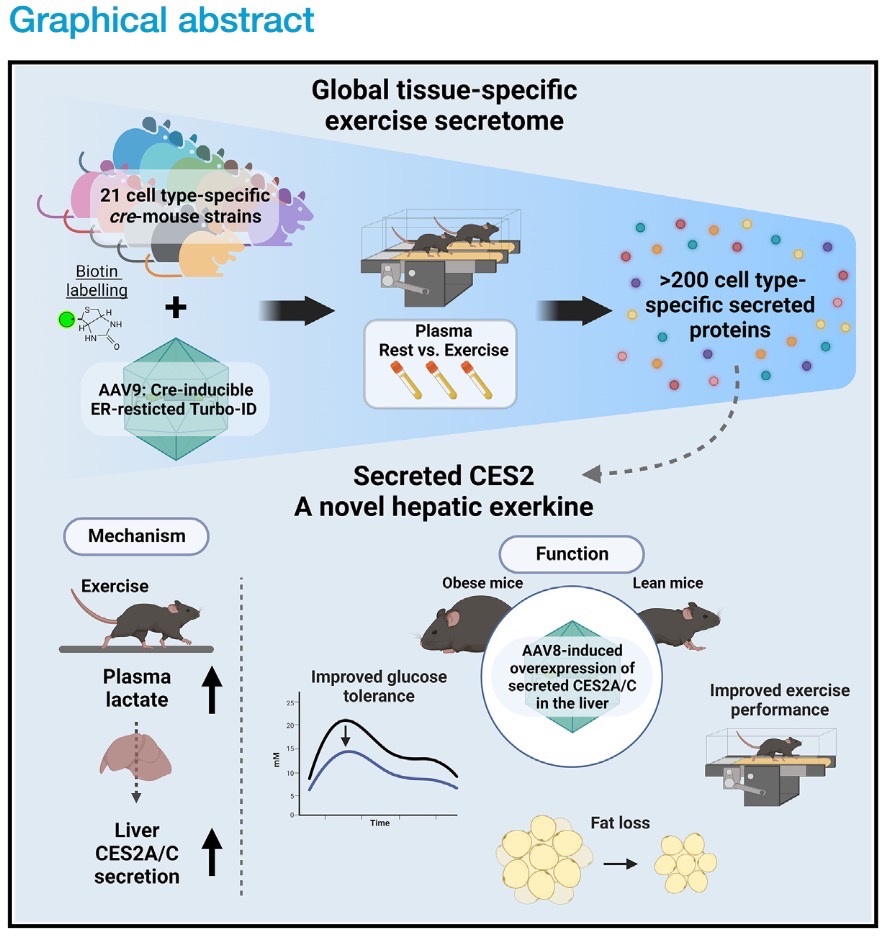
由于检测血液代谢组方法的局限性,大多研究主要关注运动对单一分子、细胞类型以及组织的影响。但是人体具有分泌功能的细胞类型很多,同一类细胞分泌不同物质发挥不同功能,不同类型细胞所分泌的同一物质也可能发挥不同的功能。因此,检测运动对多器官和细胞分泌组的影响十分重要。斯坦福大学Jonathan Z. Long课题组就这个问题进行了深入的研究【Wei … Long. Organism-wide, cell-type-specific secretome mapping of exercise training in mice. Cell Metab. (2023)】。
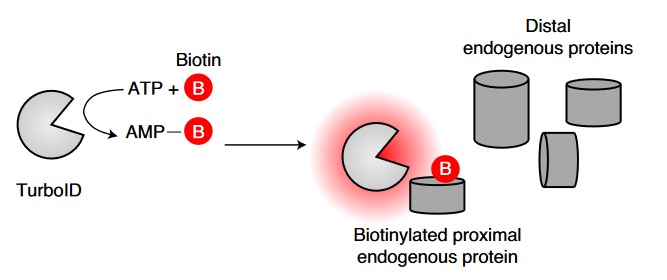
图 1 TurboID邻位标记原理
为了研究运动前后小鼠不同器官和细胞分泌组的变化,作者使用了一个标记和富集分泌蛋白质的方法。2008年,Eunjoo Choi-Rhee突变大肠杆菌生物素连接酶BirA,使其可以非特异性生物素化蛋白质,实现了生物素介导的邻位标记。此后经过一系列的人工进化,Alice Y Ting课题组获得了快速(10 min)、小范围(<10 nm)催化赖氨酸残基生物素化的生物素连接酶——TurboID。利用链霉亲和素富集纯化后的生物素标记蛋白质可以通过质谱鉴定蛋白质的种类。为了标记内质网蛋白,Jonathan Z. Long课题组在前期的研究中将内质网定位信号KDEL插入到TurboID C端构建ER-TurboID,用以标记经内质网的分泌蛋白。为了在小鼠特定细胞系表达ER-TurboID,他们将ER-TurboID以DIO(double-floxed inverted open reading frame)的形式插入到AAV9中构建了AAV9-ER-TurboID实现了Cre依赖的内质网TurboID表达。DIO含有两组反向交替排列的loxp位点,中间是倒置的ER-TurboID,Cre识别两组loxp位点后两次切割使倒置的ER-TurboID翻转以正常表达。
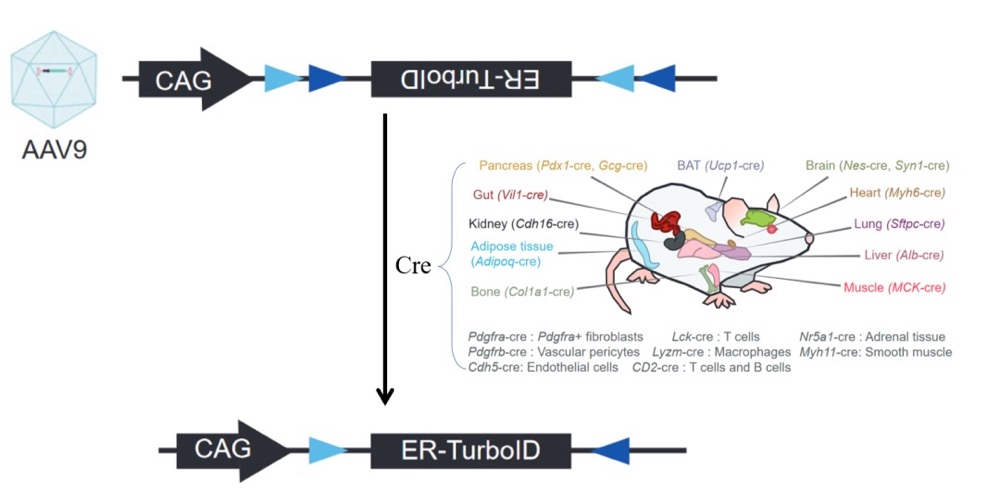
图 2 Cre依赖的内质网表达TurboID
作者分别选用了21种不同组织特异性Cre成年小鼠(例如骨骼肌、脂肪细胞、肝细胞、心脏等),采用尾静脉注射AAV9-ER-TurboID的方法分别在各组织细胞内质网中表达了TurboID。病毒注射三周后,小鼠进行每天1小时,20 m/min的跑步机训练,持续一周。在训练第四天时开始通过饮用水和腹腔注射的方式补充生物素,第七天训练结束后颌静脉取血,利用链霉亲和素对血浆中的生物素化蛋白纯化后,检测蛋白质的种类。为了确定跑步机训练实现小鼠运动的目的,在小鼠七天跑步机训练后,作者检测到已知的和运动相关基因表达量上调(Pgc1a、Nr41a)、脂肪重量减少、脂肪细胞面积减小,但是摄食基本不变。作者共检测到了1272个细胞类型-蛋白对(每种细胞所分泌的每种蛋白都视为一个独立的样本),181个变化的分泌蛋白质。其中肝细胞和肌肉细胞分泌蛋白种类最多,但是运动前后脂肪肝细胞(Pdgfra+)分泌蛋白变化数量最多。。
经过7天训练,分泌量仅在一种细胞组织中发生变化的蛋白有129种,分泌量在多种细胞组织中发生变化的蛋白有52种。例如LOXL1在所有细胞类型中分泌量都下调;EMIL1在所有细胞类型中分泌量都上调;SOD3和HSP7C存在双向调节,在一些细胞类型中上调,另一些细胞类型中下调。这些数据中作者发现了很多以前未发现的运动调节的分泌蛋白,并对这些分泌蛋白的细胞来源提供了依据。
由于运动前后分泌量蛋白变化数量最多的细胞是脂肪干细胞,作者对运动诱导的脂肪干细胞分泌组进行了GO分析。发现这些变化的分泌蛋白主要参与了“刺激响应”。为了探究这些运动诱导的分泌蛋白是否和运动持续时间相关,作者分别设置了不运动、运动1天、运动3天以及运动7天的Pdgfra-Cre小鼠,检测脂肪干细胞分泌蛋白中F13A、C4BPA和ITIH2的表达量。发现ITIH2在运动后第1天分泌量就增加并持续稳定;C4BPA分泌量在运动1-3天降低,第7天又增加;F13A分泌量在第1天增加,第3天降低,第7天又增加。说明脂肪干细胞对运动的响应具有多样性,并且部分蛋白的分泌依赖于运动时间。
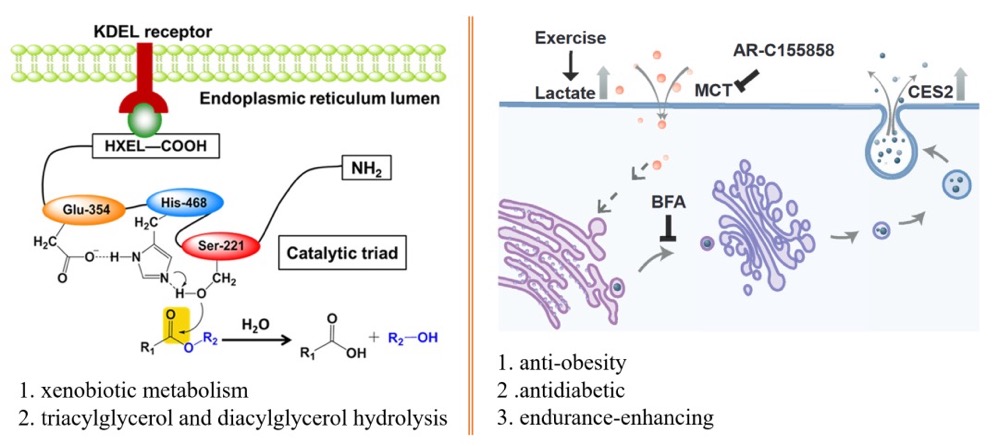
在对肝细胞分泌组进行分析时,作者发现了一组主要催化酯键水解的同工酶CES2A/C在运动后上调。CES2主要在肝脏细胞表达,通过N端HXEL序列结合KDEL序列定位于内质网。CES2被认为在含酯异生物质代谢中发挥重要作用,例如抗抑郁药物伊立替康、阿片类药物等,同时还能水解甘油三酯和甘油二酯促进脂肪代谢。文献显示肝脏细胞过表达CES2可以降低小鼠体重,但是作者在血浆中检测到了肝细胞分泌的CES2,因此作者想知道这些肝细胞分泌到血液中的CES2发挥什么样的作用。
首先作者想知道被认为定位于肝细胞内质网的CES2如何在运动后分泌到胞外。运动后肌肉产生大量的乳酸,但是无法利用,大量乳酸分泌到胞外,通过血液运输到肝脏细胞,再由肝细胞通过糖异生合成葡萄糖后运输到肌肉组织供其利用。作者想知道乳酸是否会促进肝细胞CES2的分泌,于是做了一个体外细胞实验,发现乳酸进入肝细胞后促进CES2通过经典的内质网-高尔基体-囊泡分泌途径运输到胞外。
接着作者想知道肝细胞分泌到胞外的CES2是否以及如何参与降低体重的过程。他们构建了CES2 HXEL序列突变病毒AAV8-Tbg-CES2-ΔC,并将其注射到小鼠体内,使CES2在肝细胞特异启动子Tbg调控下表达并分泌胞外。发现在肝细胞过表达分泌性CES2A/C后小鼠体重下降、血糖浓度更低、白色脂肪减少,但是其他组织没有明显变化。于是作者检测了代谢变化,发现肝细胞过表达分泌性CES2A/C后小鼠氧气消耗量增加,且CES2A可以促进小鼠的运动。作者还通过突变掉酶活性位点证明了肝细胞分泌到胞外的CES2的整体作用需要其羧酸酯酶活性,非靶向代谢组学分析提示CES2C可能更倾向于通过脂质代谢促进体重降低。
总之,本文利用定位于内质网的TurboID研究了多种组织和细胞分泌蛋白在运动后的变化,发现了很多新的响应运动的细胞类型分泌蛋白。他们发现运动产生的乳酸进入肝细胞,协助CES2A和CES2C分泌到胞外,并且探索胞外CES2在降低肥胖中的潜在机制(抗肥胖、增强葡萄糖耐受等),为日后研究细胞-细胞间相互作用提供了新思路。
运动对机体的影响显而易见,究竟是通过什么机制导致机体变化十分不清楚。虽然作者做了些探索,还有许多问题没有解决。
首先,肌肉应该是引起机体变化的主要器官,作者只是聚焦在乳酸(如对CES2的分泌)这个经典、已知的分子。
第二,运动后,细胞不仅仅分泌蛋白,可能还分泌非蛋白物质;而且分泌蛋白的方式有多种,包括直接跨膜、ABC转运体、囊泡转运以及跨高尔基转运,由于生物素定位于内质网且只能标记经由内质网分泌的蛋白质,因此该方法无法研究不经过内质网的细胞分泌物质,也无法研究非蛋白类细胞分泌物质。
第三,作者聚焦的是运动七天后分泌组的变化,变化的蛋白代表稳态的结果,包括许多细胞的适应反应,而不是组织细胞在运动后最初分泌组(即起始信号)。
第四,虽然AAV9能感染多种组织,但不同组织的感染效率不一致,单一血清型病毒的感染特性可能会影响部分细胞中TurboID的表达效率,从而使得鉴定蛋白数量偏少。
最后,作者发现乳酸促进肝细胞分泌CES2,但是对CES2整体蛋白水平没有影响,说明新产生的CES2立即分泌出细胞了,这个机制是什么?究竟血液中CES2如何调节代谢的机制仍不清楚。
尽管如此,作者通过研究多组织、多细胞的内质网分泌组在运动后的变化,为日后的机制研究提供了新思路。


 北京市丰台区右安门外西头条10号
北京市丰台区右安门外西头条10号
 010-86738999
010-86738999
 cimr@cimrbj.ac.cn
cimr@cimrbj.ac.cn









 En
En
