撰文|吴桐(2023级博士研究生)
审核|李国民
有 70 种神经退行性疾病是由基因特异性串联重复序列的扩增引起的,其中有不少与(CAG)、(CTG)重复序列的扩增有关,这包括亨廷顿病(HD)和脊髓小脑共济失调(SCA)。单链 DNA ( single-stranded DNA,ssDNA )在转录、复制、修复或重组后重新连接时,由于碱基配对错误而形成slipped-CAG DNA结构。单链结合蛋白(SSB,single strand DNA-binding protein)是生命所必需的,普遍表达,几乎介导了所有涉及ssDNA的过程,包括DNA双链的不稳定性、异常DNA结构的解链、DNA的重新接合、ssDNA的保护以及DNA蛋白的招募。人类的主要SSB是经典的 RPA(复制蛋白 A),它是由 RPA1、RPA2 和 RPA3 组成的复合物;灵长类动物还表达另一种形式的RPA,即Alt-RPA,它与经典RPA的区别在于RPA2与RPA4的对映体互换,RPA4与RPA2具有相似的结构域,而RPA4的功能尚不清楚。
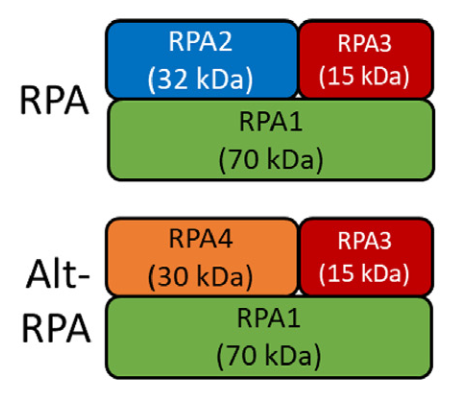
图1:经典RPA ( RPA1、2、3)与Alt - RPA ( RPA1、4、3)
2023年10月,来自多伦多大学的Christopher Pearson课题组在Cell上发表题为“Antagonistic roles of canonical and alternative-RPA in disease-associated tandem CAG repeat instability”的文章,他们发现,在亨廷顿病和脊髓小脑共济失调 1 型(SCA1)患者大脑中, Alt-RPA浓度上调,在正常情况下RPA能增强体外解链、FAN1切除和slipped-CAG的修复,并能防止人类细胞中的CAG重复序列扩增。在SCA1小鼠大脑中过表达RPA可消除扩增,同时减少ATXN1聚集、减少大脑DNA损伤、改善神经元形态并修复运动表型。相反,Alt-RPA可抑制解链、FAN1 切除和slipped-CAG的修复,并促进CAG重复序列扩增。这些发现表明这两种RPA之间存在功能性相互作用,Alt-RPA可拮抗RPA,从而加强对疾病相关CAG重复序列的扩增作用,并可扩展到其他DNA过程。

为了研究这一差异,研究人员首先对HD和SCA1患者死后大脑中RPA亚基的RNA和蛋白质水平进行了定量分析,发现在纹状体内和小脑,与未受影响的个体相比,HD 患者的所有 RPA 转录物均显著上调。
接下来他们构建了两种环状(CAG)50·(CTG)30 slipped-DNA底物,进行体外DNA修复实验,两种底物分别是在重复序列(nick-in-repeat)或侧翼序列(nick-in-flank)中带有缺口,实验发现nick-in-repeat会产生一种迁移缓慢的结构构象,这是slipped-DNA构相改变所导致的,并且加入RPA和Alt-RPA后这种中间体减弱了,研究结果表明,RPA和Alt-RPA都不是形成改变的slipped-DNA构象或修复slipped-DNA的必要条件。然而,以浓度敏感的方式,RPA会竞争性地减少改变的构象并增强slip-out修复,而高水平的Alt-RPA会竞争性地逐渐抑制slip-out修复。
为了了解修复效率的差异,研究人员通过电泳迁移试验(EMSA),使用放射性标记的线性化nick-in-repeat slipped-CAG底物,研究RPA和Alt-RPA的DNA结合情况,发现RPA和Alt-RPA均与DNA结合, RPA产生两种不同的迁移较慢的蛋白- DNA复合物,Alt-RPA只产生一个蛋白- DNA复合物,且Alt-RPA结合和解链部分双链DNA结构都比经典RPA慢。有趣的是,RPA能够增强FAN1核酸内切酶活性,而Alt-RPA会抑制FAN1核酸内切酶活性。
为了鉴定RPA和Alt-RPA的蛋白-蛋白关联,研究人员使用BioID邻近标记实验鉴定RPA1-4的近端关联蛋白,结果显示互作组相当广泛,包括:CAG不稳定修饰因子( MSH2、MSH3、MLH1、PMS1、XPG等);HD/SCA疾病修饰因子( HTT、RM2B/p53R2、UBR5、TCERG1/CA150、SETD2/HYPB/KMT3A等); CAG/CTG疾病发病机制相关;其他重复扩展病相关蛋白(RFC1、ATXN2、ATXN10、EIF4A3、DIP2B等); DNA损伤应答( P53、MRE11、NBN、LIG3、BRCA1、BRCA2、XPC等);染色质生物学;DNA/RNA代谢;DNA损伤等相关蛋白。且使用纯化的RPA和Alt-RPA复合物与MutSα(MSH2-MSH6)和MutSβ(MSH2-MSH3)蛋白复合物进行免疫共沉淀实验也支持BioID结果,即Alt -RPA与MutSβ相互作用,RPA与MutSα、MutL和HTT相互作用。而在过表达了人RPA2或RPA4的HT1080细胞模型中,发现RPA上调抑制体细胞CAG重复序列的扩增,Alt-RPA上调促进体细胞CAG重复序列的扩增。
为了评估RPA调节的体内效应,研究人员在突变型 Ataxin-1 Q135 knock in SCA1小鼠的大脑中过表达了RPA1,这些小鼠反映了 SCA1 患者的疾病特征,发现RPA上调可以降低基因组范围的DNA损伤和突变型ATXN1累积。
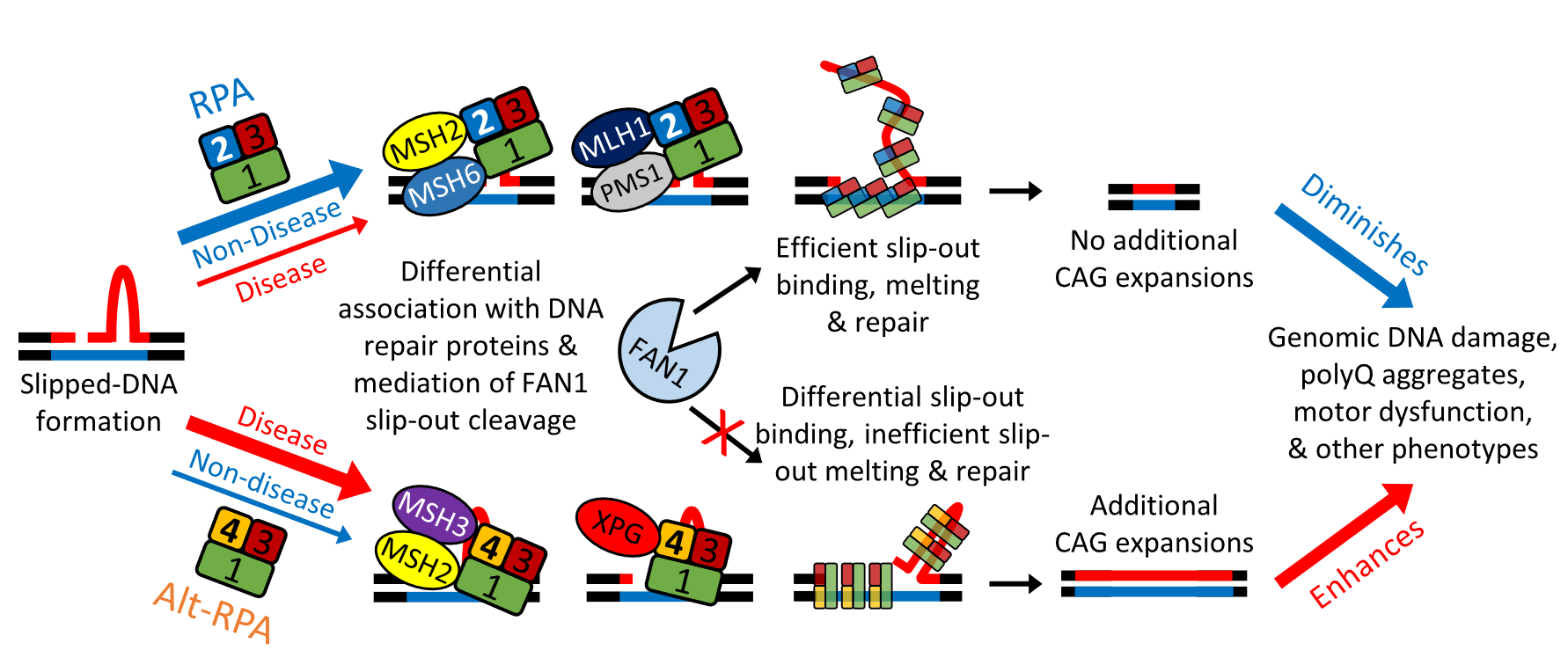
图2:Alt – RPA与RPA拮抗互作在体细胞CAG重复序列扩增中的工作模式
以上结论揭示RPA防止体细胞CAG重复序列扩增,从而减少疾病表型,而Alt-RPA减少RPA的活性,促进扩增和恶化表型,模型如图2。重复扩增突变涉及单链DNA中间体和异常DNA结构,需要SSB来稳定、保护、解链和退火单链,并招募DNA修复蛋白。经典RPA可通过快速解链slip-outs、加强FAN1介导的slip-outs切除、减少CAG重复序列扩增和减少下游事件来提高正确修复率;Alt-RPA抑制修复、低效解链slip-out和抑制FAN1介导的切除,从而导致扩增并加剧下游事件(本研究显示的下游事件包括脑DNA 损伤和神经元多聚谷氨酰胺聚集体、运动表型、神经元形态以及转录组、剪接体和细胞周期失调)。在非疾病状态下,上层途径可能占主导地位,而在疾病状态下(RPA/Alt-RPA 水平失调),下层途径可能占主导地位。RPA/Alt-RPA 与已知可调节重复扩增的DNA修复蛋白的不同关联也会导致不稳定性。RPA优先与 MutSα(MSH2-MSH6)和MutLβ(MLH1-PMS1)相互作用。Alt-RPA 优先与 MutSβ(MSH2-MSH3)和 XPG 相互作用。
总的来说,这项研究揭示了单链DNA结合蛋白RPA和Alt-RPA在表达、功能和蛋白-蛋白互作上的平衡调控与神经退行性疾病相关的体细胞CAG重复序列扩增,对于干预相关神经退行性疾病具有重要意义。
但文章仍存在不足,主要问题在于:
1. 在突变型 Ataxin-1 Q135 knock in SCA1小鼠的大脑中过表达了RPA1后,为什么RPA1的过表达会同时引起RPA2、RPA3的表达上调?
2. 经典RPA与Alt-RPA差异在于异源三聚体组成不同,为什么不在小鼠模型里分别表达RPA4和RPA2或测证是RPA4引起了CAG重复序列的扩增?



 北京市丰台区右安门外西头条10号
北京市丰台区右安门外西头条10号
 010-86738999
010-86738999
 cimr@cimrbj.ac.cn
cimr@cimrbj.ac.cn









 En
En
