
撰文|冯欣然
审核|梅林

在大脑中,数十亿的神经元通过数量庞大的突触特异性地进行相互连接,然而实现这种特异性的分子机制仍然大多数情况下是未知的。对于这种偏向性连接的一个合理解释是,某些细胞类型的过程会表达细胞识别蛋白,这些蛋白会选择性地结合到伙伴细胞表面上的受体上,这种识别事件可能会触发促进适当伙伴之间突触形成的粘附相互作用[1]。
2019年,斯坦福大学Thomas C. Südhof团队发表了一篇《科学》文章[2],并报告称小鼠海马CA1区域的突触形成需要latrophilin 2和3(Lphn2和Lphn3)。有趣的是,Lphn2介导自内嗅皮层投射的突触形成,而Lphn3介导自CA3区域投射的突触形成。这一功能需要突触后的latrophilin 与突触前的teneurins和FLRTs之间的结合。
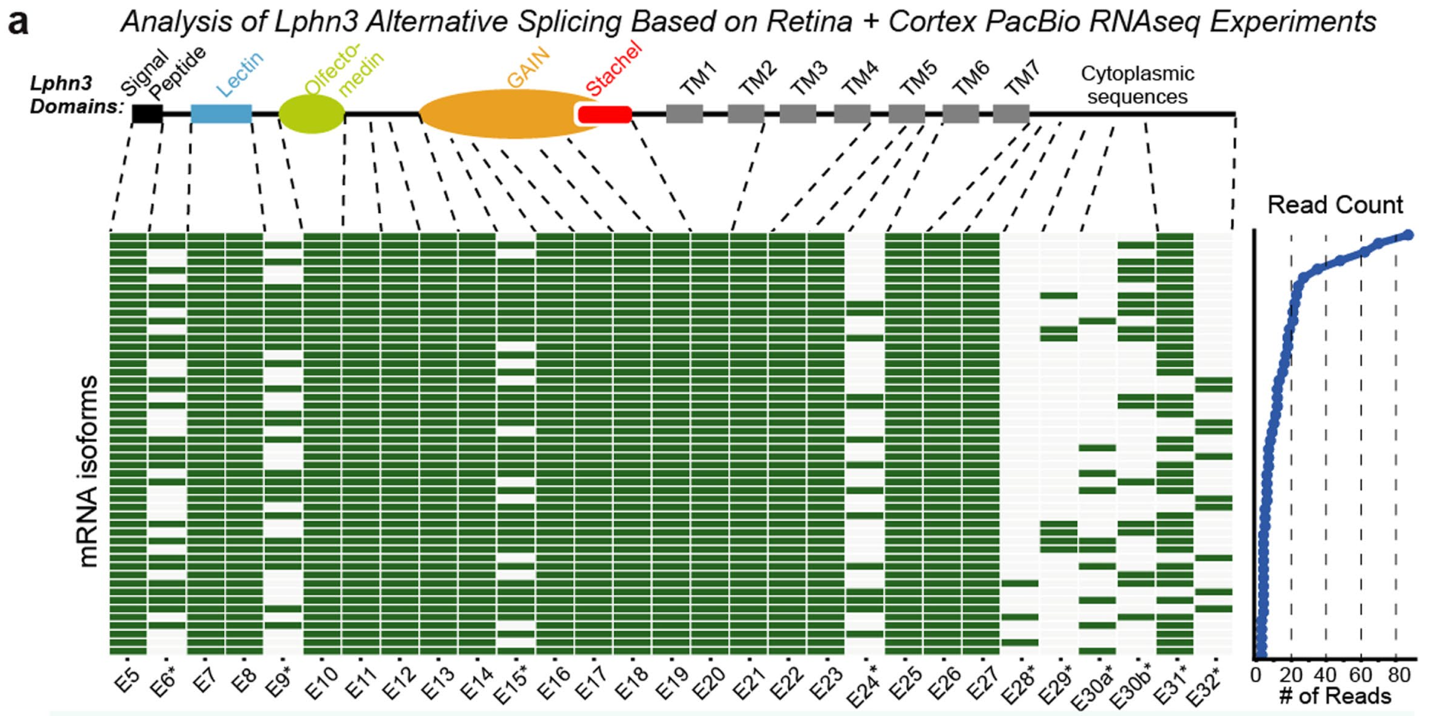
Lphn3是一个由众多外显子编码的多结构域蛋白质。在一个称为“RNA剪接”的过程中,从基因组转录出的前体mRNA在被运送到细胞质进行蛋白翻译之前,被剪接因子进一步处理,将内含子切除,而保留外显子部分。然而,在大多数情况下,一个基因可以经历不同的剪接模式,即所谓的可变剪接(AS)。可变剪接可能导致mRNA降解或产生多样的蛋白异构体。通过分析PacBio RNA-seq数据和神经元亚型特异性RNA-seq数据,作者发现Lphn3有32个外显子,其中至少有10个外显子经历了可变剪接。许多选择性外显子在兴奋性和抑制性神经元中显示出独特的剪接模式,特别是外显子31(E31)和32(E32)。首先,E31在抑制性神经元中被更多地剪入。E32在兴奋性神经元中被更多地剪入。有趣的是,E31和E32实际上是互斥(mutually exclusive)的。从蛋白序列上看,E30、E31和E32的剪接确定了Lphn3的细胞质端(也是C端)结构。第三,Lphn3外显子的剪接模式在大脑区域和出生后发育阶段也有所不同。其中,包含E31的变体是小鼠海马中丰度最高的Lphn3 C端剪接变体。
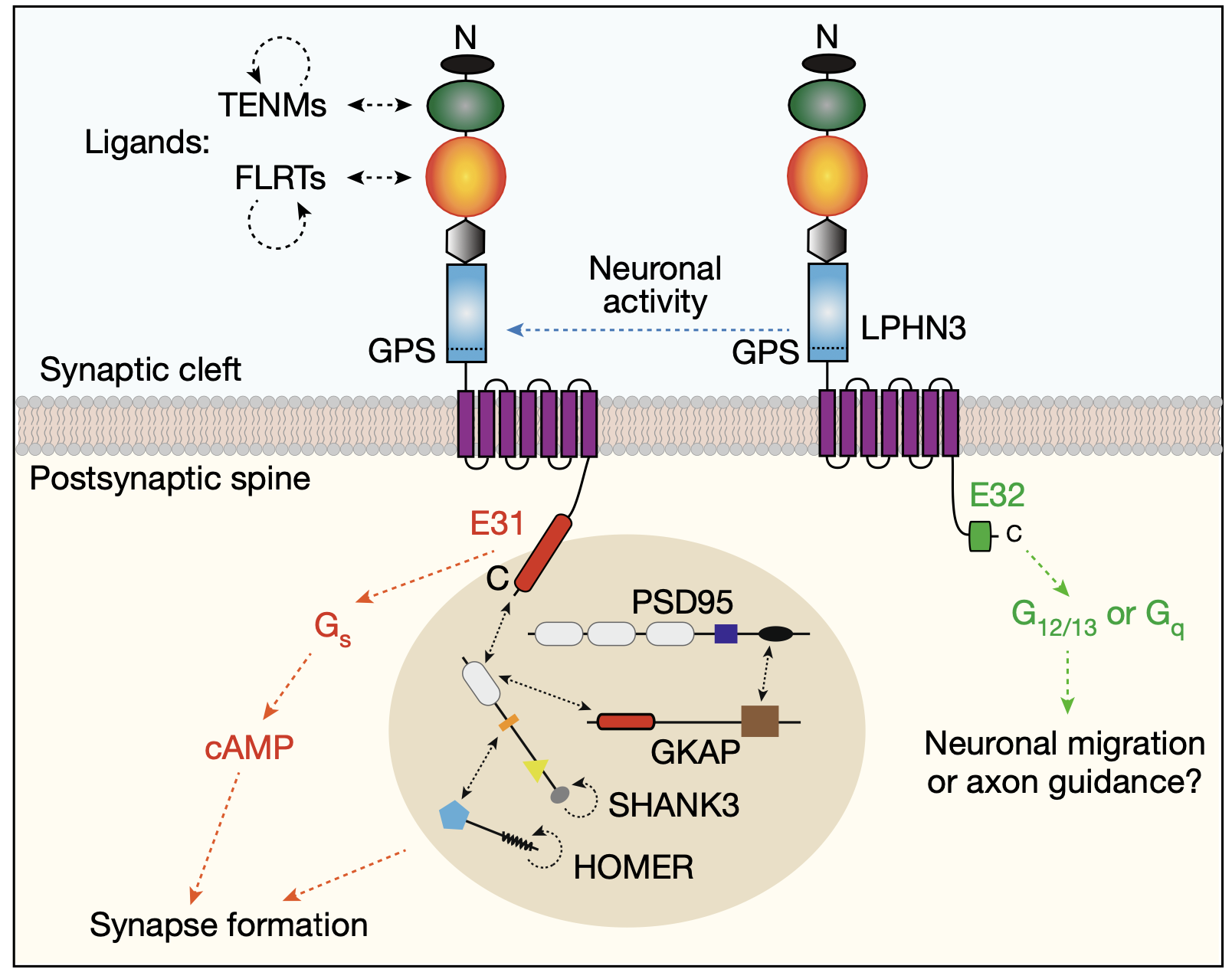
考虑到GPCR的C端对于G蛋白耦合功能至关重要,作者随后探究了Lphn3 C端剪接事件是否改变其对Gα蛋白偶联的偏好。他们利用TRUPATH,一个用于探究GPCR转导体组的开源生物传感器平台,通过BRET2(生物发光共振能量转移)实验来衡量Lphn3各异构体的G蛋白偶联能力。他们发现E31变体更倾向于与Gαs结合,而E32变体更倾向于与Gα12/13结合。E31变体的过表达显著增加了细胞中的cAMP水平。有趣的是,他们还注意到,Lphn3的同源基因Lphn1和Lphn2的剪接模式也可能调节它们的G蛋白偶联能力。
所以究竟哪种Lphn3 C端剪接变体促进了突触形成?为了回答这个问题,作者使用CRISPR敲除了含有E31的Lphn3转录本。E31 KO并没有改变Lphn3蛋白水平,但相反增加了含有E32的转录本水平。他们发现,E31特异性KO降低了神经元的放电频率,这与全局性敲除Lphn3蛋白所观察到的情况相符。同时,E31特异性KO,就像全局KO一样,减少了兴奋性突触的密度。
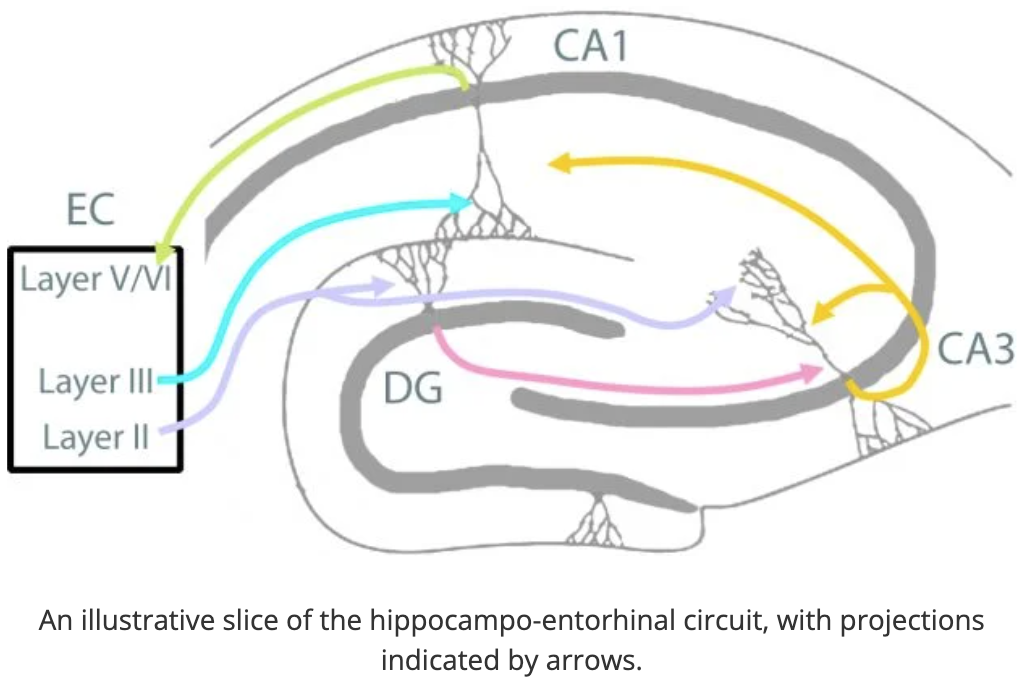
如上所示,海马CA1神经元主要接收来自内嗅皮层(EC)和海马CA3区域的神经输入[3]。Südhof及其同事此前表明,海马CA1区域的Lphn3介导了来自CA3区域而非内嗅皮层的突触形成。为了研究E31特异性KO是否会影响这一功能,作者使用单突触逆行示踪技术追踪到CA1区域的神经连接。他们发现,E31特异性KO减少了来自同侧和对侧CA3区域对CA1金字塔神经元的突触输入。因此,与Gαs结合的含E31的异构体对于Lphn3介导的突触连接至关重要。
接下来,作者探究了Lphn3 E31剪接变体是否除了与Gα蛋白偶联外还具有其他功能?含有E31的Lphn3异构体在C端具有一个PDZ结合基序(PBM),而含有E32的Lphn3异构体则没有。这个基序与后突触后蛋白组装体中的Shank蛋白相互作用。因此,作者进行了包括GKAP、Homer、PSD95、Shank(GHPS,四个共同发生相分离的突触后支架蛋白)以及一系列Lphn3变体和突变体在内的体外沉淀实验。如预期的那样,只有含有E31的Lphn3与GHPS复合物共沉淀,而含有E32或E31(ΔPBM)的Lphn3没有。将前突触配体Lphn3、TENM2和FLRT3添加到混合物中促进了相分离液滴的聚集。同样,这种效应在添加含有E32的Lphn3或不含PBM的E31时未被观察到。值得注意的是,Lphn3表现出比大多数支架蛋白更快的恢复动力学,这表明Lphn3 E31变体在后突触支架蛋白凝聚体表面形成了流动的壳层。因此,LPHN3 E31而非LPHN3 E32会定位于由突触后支架蛋白形成的相转移液滴表面,并且LPHN3的突触前配体TENM2和FLRT3可以进一步使这些液滴聚集。
那么,Lphn3 E31编码的PDZ结合基序是否对突触形成至关重要?为了回答这个问题,作者再次使用CRISPR有选择性地在含有E31的Lphn3变体中删除了PBM基序。这种操作并没有减少蛋白质或mRNA水平,也没有改变Lphn3的剪接模式。然而,正如预期的那样,PBM的删除显著降低了海马神经元中的兴奋性突触密度。因此,Lphn3 E31编码的PBM对兴奋性突触的形成是至关重要的。
最终,作者们探究了Lphn3 E31和E32的剪接是否受神经元活动调控,因为这两个外显子在不同类型的神经元和突触中显示出明显不同的模式。作者分析了已发表的RNA-seq数据集,发现当使用氯化钾、PTX或KA激活神经元时,Lphn3的剪接模式从含有E32转变为E31。此外,单细胞RNA-seq表明,具有更高即早基因(immediate early gene)表达的神经元也具有更高的E31剪入比率。
总体而言,这篇论文架构良好,实验设计详实。然而,我对作者仍有一些疑虑和问题:
原文献:
Wang, S., DeLeon, C., Sun, W., Quake, S.R., Roth, B.L., and Sudhof, T.C. (2024). Alternative splicing of latrophilin-3 controls synapse formation. Nature 626, 128-135.
参考文献:
[1] Sanes, J.R., and Zipursky, S.L. (2020). Synaptic Specificity, Recognition Molecules, and Assembly of Neural Circuits. Cell 181, 536-556.
[2] Sando, R., Jiang, X., and Sudhof, T.C. (2019). Latrophilin GPCRs direct synapse specificity by coincident binding of FLRTs and teneurins. Science 363.
[3] Petrantonakis, P.C., and Poirazi, P. (2014). A compressed sensing perspective of hippocampal function. Front Syst Neurosci 8, 141.
[4] Wallis, D., Hill, D.S., Mendez, I.A., Abbott, L.C., Finnell, R.H., Wellman, P.J., and Setlow, B. (2012). Initial characterization of mice null for Lphn3, a gene implicated in ADHD and addiction. Brain Res 1463, 85-92.


 北京市丰台区右安门外西头条10号
北京市丰台区右安门外西头条10号
 010-86738999
010-86738999
 cimr@cimrbj.ac.cn
cimr@cimrbj.ac.cn









 En
En
