
撰文 | 邹林君(2024级博士研究生)
审核 | 任会霞
背景介绍
结直肠癌癌症(CRC)表现出B细胞浸润和/或三级淋巴结构(TLS)形成,肿瘤浸润B细胞的功能特性及其对免疫的影响仍存在争议。由于其抗原呈递和抗体产生功能,B细胞具有抗肿瘤的免疫能力,显示出与改善癌症结果的积极关联,特别是当发现它们与TLS相关时。多种小鼠肿瘤模型表明,存在免疫抑制或调节性B(Breg)细胞亚群,这些亚群可以迁移到肿瘤床中并获得调节表型,例如抑制分子的高表达,包括程序性死亡配体-1(PD-L1)、白细胞介素-10(IL-10)和转化生长因子-β1(TGF-β1),从而导致局部抗肿瘤免疫反应的减弱。进一步剖析肿瘤环境(TME)中B细胞的异质表型和获得调节功能的机制,可能为调节CRC的免疫反应和有意义的治疗干预提供重要途径。
营养物质驱动细胞代谢并决定细胞命运,然而,TME缺乏营养,尤其是对葡萄糖的竞争和消耗。低血糖限制了TME中细胞毒性免疫细胞和调节性T(Treg)细胞的分化和功能。这些细胞利用脂质、氨基酸,甚至肿瘤代谢产物,如乳酸,来获取能量并维持其生存和功能。氨基酸调节肿瘤细胞的增殖驱动,并作为蛋白质合成底物发挥直接作用。氨基酸也在能量代谢、维持细胞氧化还原稳态和表观遗传修饰中发挥作用。之前有报道已经证明了通过RagA蛋白进行氨基酸传感在协调胚胎发育、介导造血干细胞功能和促进Treg细胞功能中不可或缺的作用。然而,没有证据表明氨基酸与肿瘤浸润B细胞的功能异质性有关。
亮氨酸(Leu)是一种必需氨基酸,是维持肌肉细胞功能的重要能量来源,通常作为恶性肿瘤患者的营养补充剂。该研究发现在CRC中富集的Breg细胞高度表达线粒体亮氨酸-tRNA合成酶-2(LARS2),显示出对亮氨酸的营养偏好。LARS2表达的B细胞(LARS B)具有TGF-β1显性调控表型,并由于LARS2的高表达而显示出增强的线粒体蛋白生物合成。亮氨酸通过促进还原的NAD+(NADH)脱氢酶的LARS2依赖性翻译来促进线粒体烟酰胺腺嘌呤二核苷酸(NAD+)再生,从而编程氧化代谢并促进TGF-β1的产生。LARS B细胞缺失抑制TGF-β1介导的免疫抑制和CRC生长。在CRC患者中,LARS B细胞位于TLS外,与肿瘤生长加速有关。肿瘤床中LARS B细胞的频率与肿瘤增生和患者生存期缩短呈正相关。在最后,对于CRC治疗,研究人员提出了一种亮氨酸节食方案来阻断B细胞介导的免疫抑制。
Graphical Abstract
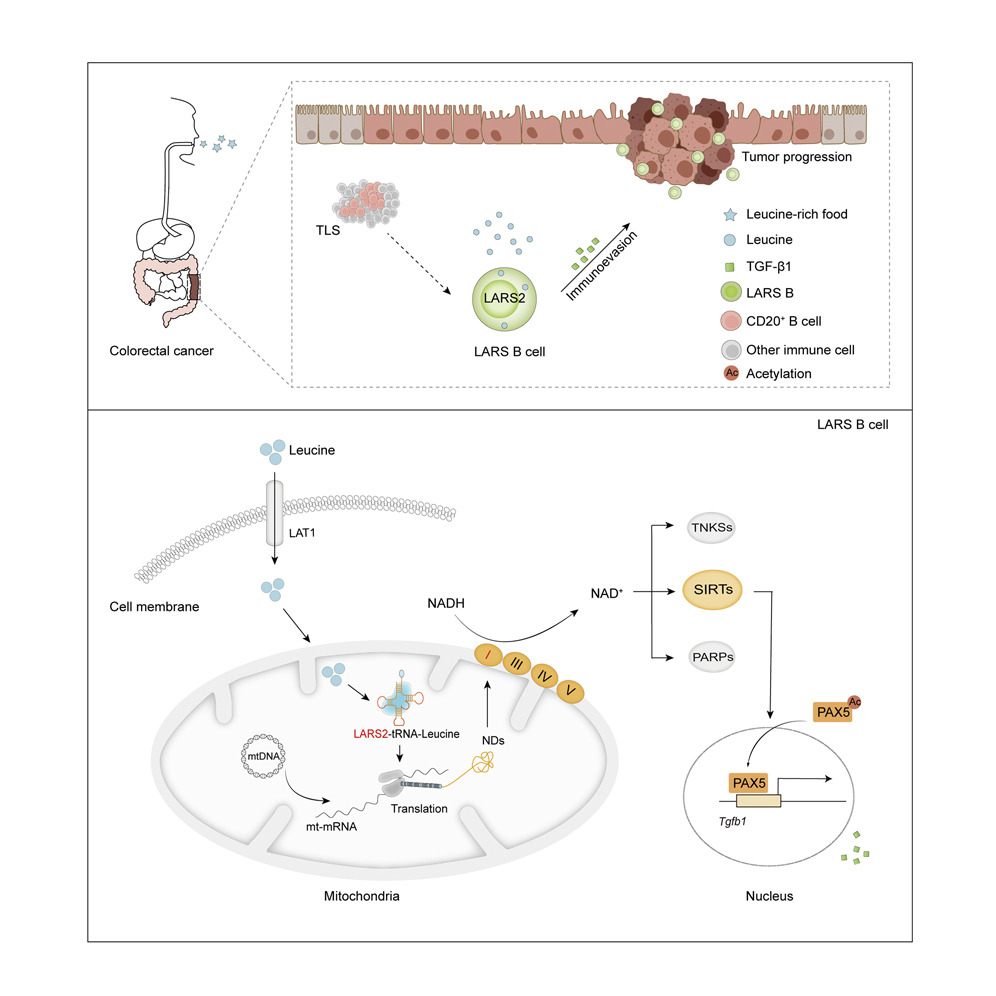
研究结论
1. 具有TGF-β1主导调控特征的LARS B细胞亚群存在于进行性结直肠癌癌症中
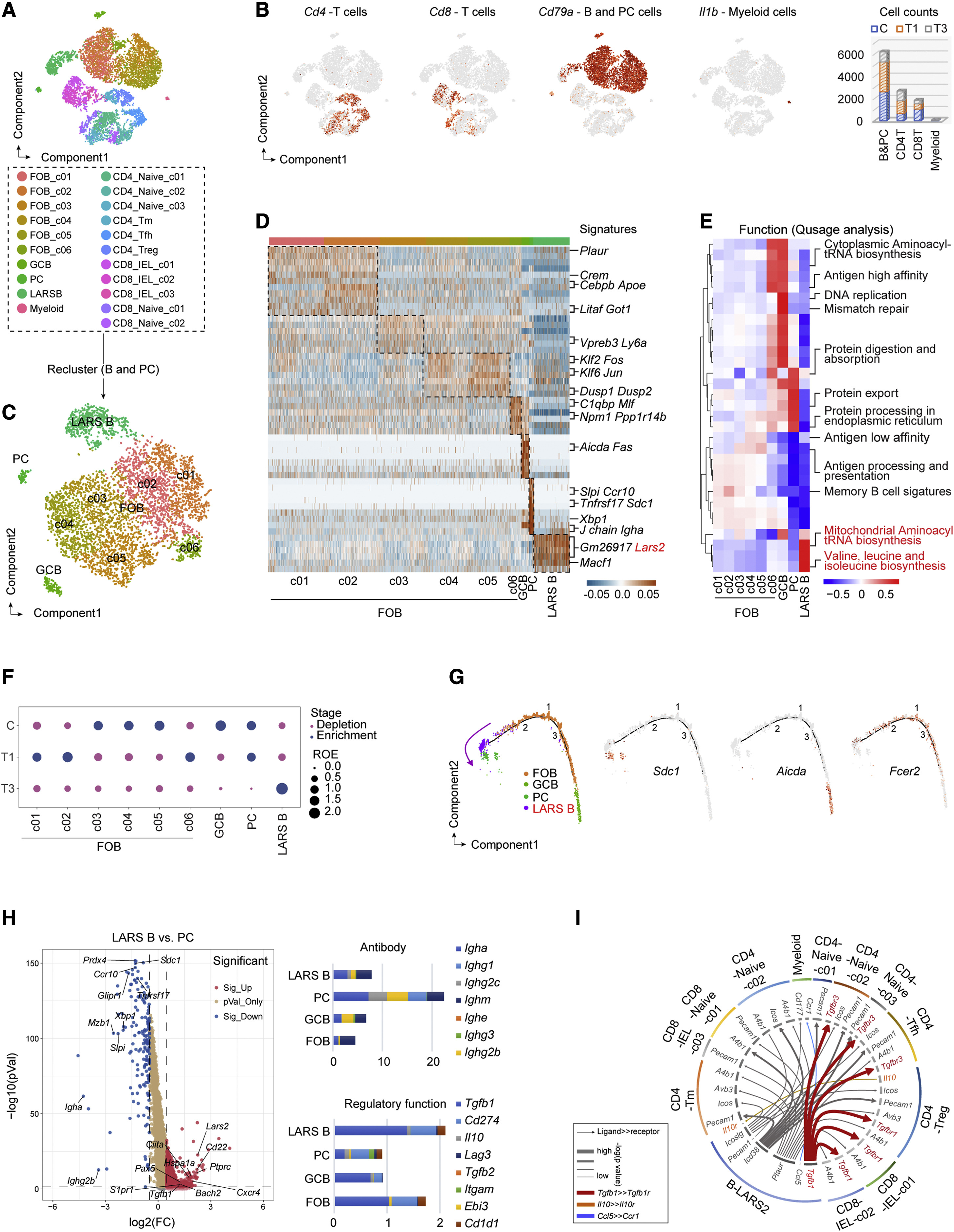
Figure 1. The LARS B cell subset is detected in progressive colorectal cancer by single-cell sequencing
为了对伴随肿瘤生长的各种B细胞亚群进行分类,在偶氮氧甲烷(AOM)+葡聚糖硫酸钠(DSS)小鼠CRC模型下,对低和高级别上皮内瘤变(T1和T3)组织的局部CD45+免疫细胞进行了单细胞测序(scRNA)分析。在细胞聚类后,对体B细胞和浆细胞(PC)进行重新聚类,LARS B细胞亚群被鉴定为具有高表达的线粒体亮氨酸tRNA合成酶-2(LARS2)显示出亮氨酸生物合成和线粒体氨酰tRNA生物合成的增强功能。此外,随着肿瘤的生长,LARS B细胞的数量与其他B细胞相比有所增加。此外,细胞通讯分析显示,LARS B细胞与IL-10+FOXP3+Treg细胞之间存在密切的相互作用,主要取决于LARS B淋巴细胞中TGF-β1的高表达。
2. LARS B细胞位于TLS外,与结直肠癌癌症患者生存期缩短有关
研究者通过免疫组织化学染色观察了小鼠和人类LARS B细胞的位置和组织特异性。
与大多数在TLS中收获的CD19+B细胞(存在于癌症与正常组织的交界处)相比,LARS B细胞主要位于TLS外部并分散在肿瘤间质中。正如预期的那样,他们还使用组织微阵列和TCGA数据库分析了LARS B细胞与预后生存之间的关系。这些结果表明LARS B细胞促进CRC进展。
3. LARS B细胞是亮氨酸诱导的,有助于结直肠癌癌症免疫逃避
为了探索LARS B细胞的产生,研究者检测了CRC浸润B细胞的各种必需氨基酸。从AOM加DSS-诱导的CRC小鼠的高级别上皮内瘤变大肠癌癌症(T3)和正常组织(WT)纯化的B细胞中的必需氨基酸。结果表明,亮氨酸是B细胞吸收最多的氨基酸。另一方面,LAT1抑制剂阻断亮氨酸向B细胞的转运会损害LARS B细胞的生成和TGF-β1的分泌。随着亮氨酸浓度的增加,流式细胞术和免疫印迹检测到LARS2同样增加。此外,TGF-β1的分泌根据掺入亮氨酸的浓度而增加。
4. LARS2通过编程NAD+依赖的线粒体氧化代谢来确定调节功能
从WT或LARS2缺失(KO)B细胞小鼠的脾脏中分离出B细胞,CD4+T细胞来自WT脾脏。将它们共培养,以确定在用丝裂霉素C灭活的MC38肿瘤细胞存在的情况下,有或没有LARS2的B细胞对CD4+T细胞分化为FOXP3+Treg细胞的影响。加入抗TGF-β1抗体(αTGF-β1)(10 μg/mL)进行中和。结果表明,B细胞中LARS2的缺失显著降低了TGF-β1诱导的Treg细胞分化。为了直接证实TGF-β1对LARS B细胞功能至关重要,从CRC小鼠的肿瘤组织中纯化了LARS B(CD45+CD19+B220-CD138-IgD-)和其他B细胞,然后与CD4+T细胞体外共培养,其中TGF-β1被中和或未被中和。这些实验表明,LARS B细胞诱导CD4+T细胞分化为Treg细胞,而TGF-β1的中和作用抑制了这种分化。并且LARS B细胞是亮氨酸诱导的,并以TGF-β1依赖的方式参与免疫逃避。
5. NAD+依赖性SIRT1促进LARS B细胞产生TGF-β1
为了解释LARS B细胞的调节功能,他们研究了线粒体功能和能量代谢,因为LARS2位于线粒体中,负责线粒体基因翻译,包括线粒体呼吸链复合物I-V。结果表明,在LARS2缺失的B细胞中,线粒体呼吸链复合物的蛋白质表达受到抑制,特别是复合物I,NADH脱氢酶,导致NADH的NAD+再生受阻。为了确定NAD+是否以及如何增强LARS B细胞的调节功能和肿瘤逃逸,他们分别将Sirt1fl/fl和Sirt7fl/fl小鼠与Cd79a-Cre小鼠杂交,以产生B细胞特异性缺失SIRT1和SIRT7的小鼠(Cd79a-Cre-Sirt1fl/f1和Cd79a-Cre-Sirt7fl/fl)。不同的小鼠模型表明:Cd79a-Cre-Sirt1fl/fl小鼠的肿瘤大小较小,肿瘤结节较少。此外,TGF-β1在B细胞中的表达最高,在SIRT1缺陷的B细胞中表达降低。
6. 亮氨酸饮食抑制LARS B细胞介导的结直肠癌癌症免疫逃避
为了鉴定SIRT1的靶标,他们在SIRT1抑制后对B细胞中的赖氨酸(Lys)乙酰化进行了蛋白质组学定量。这些数据确定了受SIRT1影响的多种途径。具体来说,作为控制B细胞发育和分化的主要转录因子,PAX5被预测为SIRT1的靶点。从动物转录因子数据库(TFDB)和PROMO实验室预测了Tgfb1的候选转录因子,然后使用单细胞测序数据对LARS B簇中的5个候选转录因子进行了表达分析。Pax5基因也在LARS B细胞中表达,并在TGF-β1的启动子中显示出结合位点。通过共免疫沉淀(CoIP)在B细胞中证实了SIRT1和PAX5之间的相互作用。
7. 亮氨酸饮食抑制LARS B细胞介导的结直肠癌癌症免疫逃避
最后,他们专注于LARS B细胞作为CRC免疫治疗靶点的潜在应用。由于LARS B细胞是亮氨酸依赖性的,他们制定了几种亮氨酸节食方案来限制LARS B电池介导的CRC逃避。亮氨酸节律性节食是有效和安全的,尤其是“4+3”节食模式(LDD-4/3,正常饮食4天,然后是无亮氨酸饮食3天)。在“4+3”节食的小鼠中,LARS B细胞和肿瘤生长均被成功抑制,对体重和死亡率没有显著影响。
总结与讨论
这项研究结果在结直肠癌中揭示了LARS B细胞亚群与TGF-β1的显性调节特征。虽然该研究已经说明了LARS B细胞的表型和位置,但是LARS B细胞的发育轨迹及其与其他Breg亚群的差异还需要通过综合比较来揭示。此外,这些结果主要来源于对小鼠的研究。在结直肠癌患者中,LARS B细胞被发现与生存期缩短有关,而它们的调节功能还需要进一步研究。对于潜在的临床应用,也需要更多的工作来确认“4+3”亮氨酸节食方案对于控制患者的CRC是有用和安全的,其中LARS B细胞的贡献也有待确定。
参考文献
Wang, Zhiqiang et al. “Leucine-tRNA-synthase-2-expressing B cells contribute to colorectal cancer immunoevasion.” Immunity vol. 55,6 (2022): 1067-1081.e8. doi:10.1016/j.immuni.2022.04.017


 北京市丰台区右安门外西头条10号
北京市丰台区右安门外西头条10号
 010-86738999
010-86738999
 cimr@cimrbj.ac.cn
cimr@cimrbj.ac.cn









 En
En
