
撰文 | 赵琪玮(2024级博士研究生)
审核 | 张昱
背景介绍
在过去10年中,免疫治疗的发展使癌症治疗取得显著进展。免疫检查点阻断已成为多种恶性肿瘤的关键治疗方法。其中,aPD-1(antiPD-1)免疫检查点阻断剂已被证明在多种癌症中可以诱导持续的抗肿瘤免疫反应 [1]。然而,多数患者对治疗无应答,或者在最初的反应后出现疾病复发 [2]。因此,找到这部分患者无应答的原因并开发可以提高基于抗体的免疫检查点抑制剂的效力和反应率的方法是至关重要的。
随着对于肿瘤免疫反应的研究越发深入,人们越来越认识到树突状细胞(Dendritic cells,DCs)对于有效的抗肿瘤免疫和成功的aPD-1治疗至关重要。最相关的细胞亚型是传统的I型树突状细胞(cDC1s)。以往研究发现,触发T细胞DC串扰,并与aPD-1治疗相结合,可以有效地增强抗肿瘤免疫力 [3]。另有研究发现人类肿瘤组织中的DC频率与PD-1检查点封锁的阳性反应直接相关 [4]。此外,最近的数据表明,在aPD-1给药的活跃阶段也需要DCs才能对治疗产生有效反应 [5]。这些发现强调了T细胞与DC在肿瘤微环境(Tumor microenvironment,TME)中的相互作用对于引发有效的抗肿瘤免疫反应的重要性。
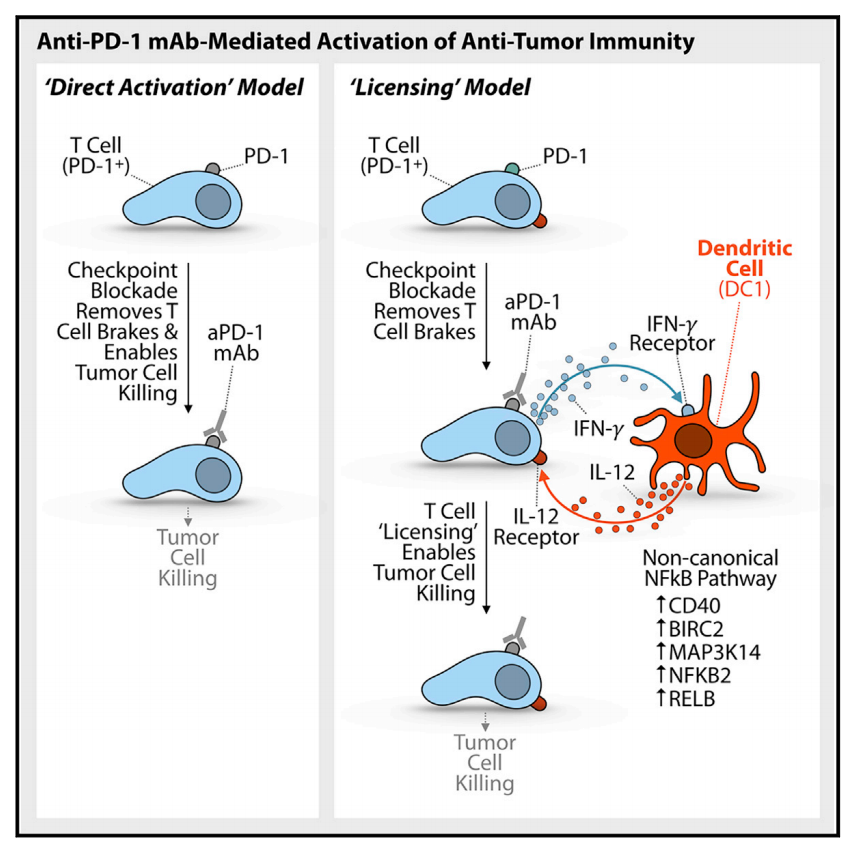
2018, Immunity 49, 1148–1161
因此,本文的作者应用了TME的单细胞转录分析接受aPD-1治疗后的肿瘤组织识别参与抗肿瘤免疫的DC相关通路,并基于此结果设计并开发了一个新的促进PD-1+ T细胞-DC物理相互作用的双特异性抗体家族以增强肿瘤特异性免疫疗法的疗效。
研究结论
1. cDC1s介导aPD-1免疫治疗中的T细胞功效
基于这些背景,研究人员设计了一系列实验来开发一种新的促进肿瘤特异性免疫免疫的疗法。首先,为了揭示aPD-1治疗期间控制CD8+ T细胞和DC串扰的分子表型,他们分析了CD8+ T细胞的动态转录流形,并追踪到9种分子层面不同的CD8+ T细胞状态。通过该图谱,作者观察到cDC1趋化因子Xcl1在TME中在效应早期T细胞和IFN-ghigh效应T细胞(与aPD-1治疗的肿瘤特异性CD8+细胞毒性反应的早期阶段相关的细胞状态)中高表达。研究人员进一步从数据中推导出了DC募集(DC-r)基因模块,发现在抗体治疗开始后的早期和晚期,aPD-1治疗的小鼠中DC-r模块的表达明显高于未治疗的小鼠。
为了进一步了解cDC1在介导aPD-1治疗作用中的作用,他们利用了Xcr1-iDTR小鼠。通过在Xcr1-iDTR小鼠中进行测试,评估存在或不存在cDC1s的情况下,aPD-1治疗对肿瘤特异性TOT-1细胞增殖和募集的影响,并发现aPD-1治疗5天后肿瘤特异性CD8+细胞出现率显著增加,但当注射DTx造成cDC1耗竭时,这种增加没有发生。此外,他们还检测了DC耗竭时aPD-1单克隆抗体单药治疗对B16-OVA肿瘤的影响,发现当cDC1在治疗开始前24小时几乎完全耗竭时,aPD-1单克隆抗体单药治疗后肿瘤生长的减少完全丧失。
上述发现表明在aPD-1治疗的活跃阶段,CD8+PD-1+ T细胞与cDC1s的相互作用对于增强CD8+PD-1+ T细胞增殖和介导有效的细胞毒性抗肿瘤反应至关重要。
2. 双特异性DC-T细胞接合剂(BiCE)
由于他们的研究结果强调了PD-1+ T细胞-cDC1相互作用的重要性,并且cDC1在TME中是一个相对罕见的免疫群体,作者假设T细胞-cDC1相互作用的低概率是T细胞对aPD-1治疗有效反应的限制因素。因此,他们设计了一种双特异性抗体(bsAb)来促进cDC1和PD-1+ T细胞之间的物理相互作用,以促进抗肿瘤免疫激活。并将该试剂称为双特异性DC-T细胞接合器(BiCE)。BiCE使用天然IgG样双特异性形式设计,其中一个Fab臂结合PD-1并阻断其与PD-L1的相互作用,另一个Fab结合cDC1表面标记CLEC9A(DNGR1)。
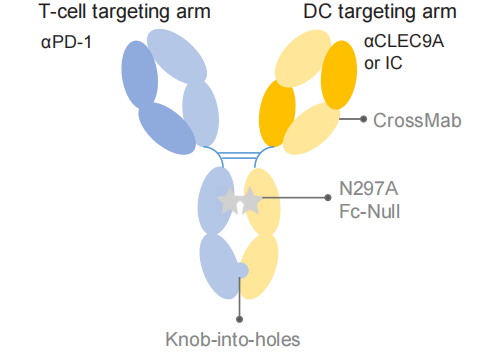
通过一系列体外实验,作者认为BiCE有效地促进了T细胞和cDC1的细胞桥接。
3. BiCE形成活化的DC-T细胞偶联物
为了评估BiCE在体内靶向形成T-DC突触的能力,作者在多个时间点分析了BiCE注射小鼠的B16F10黑色素瘤肿瘤和肿瘤dLN组织。
结果表明,BiCE在体内直接作用于 CLEC9A+cDC1与PD-1+ T细胞并增强其物理相互作用,增加了aPD-1治疗小鼠的dLN中cDC1细胞和DC/T细胞偶联物的数量。接下来,为了分析BiCE诱导的cDC1-T细胞偶联物的分子表型,他们对BiCE治疗的小鼠的dLN组织进行了物理相互作用细胞的单细胞测序技术。发现 BiCE处理诱导的物理相互作用的DC和T细胞显示了强大的免疫反应,并导致T细胞的激活和增殖以及cDC1的成熟。因此认为BiCE治疗增加了活化的DC/T细胞偶联物的丰度,从而增强了免疫功能。
接下来,作者评估了BiCE介导的DC-T细胞偶联激活对抗肿瘤免疫的作用模式。通过对小鼠dLN细胞的分析,发现在BiCE处理后,CD8+ T细胞与cDC1形成一个活跃的突触,增强CD8+ T的激活、增殖和细胞毒性活性。随后通过双态形成动力学来评估肿瘤部位的BiCE介导的T-DC相互作用。在治疗开始1天后,TME中T-DC偶联物小幅增加,但cDC1细胞的总体频率下降。这表明了BiCE并非通过提升cDC1在肿瘤中的频率促进T-DC相互作用。此外,肿瘤部位DC/T细胞串扰诱导的早期事件可能有助于cDC1的激活和向dLN的迁移,因为作者观察到在TME的T-DC偶联物在药物作用3天后进一步增加。
总的来说,BiCE治疗促进了dLN和肿瘤中活性T细胞-cDC1双链的动态形成,导致细胞毒性CD8+ T细胞的增殖和激活。
4. BiCE促进了强大的抗肿瘤活性
为了检测BiCE治疗导致的DC-T细胞相互作用的治疗效果,作者建立了三种侵袭性肿瘤模型并检测BiCE治疗、aPD-1治疗及联合治疗后的肿瘤生长情况。结果发现BiCE治疗,而不是传统的aPD-1治疗,产生了强大的抑制肿瘤增殖及转移的效果,以及远端抑制效果,表现为在注射部位和远端部位对肿瘤体积的显著控制,以及对肺转移发生率的显著控制。
基于上述结果,作者认为,BiCE无论是作为单一治疗还是作为联合治疗的一部分,都能持续诱导强大的抗肿瘤反应,并在多种侵袭性肿瘤模型中显示出治疗效果,包括对传统aPD-1治疗有耐药性的癌症类型。这种效率不仅延伸到原发肿瘤,也延伸到全身环境和抑制转移。
5. BiCE介导的串话增强了TME中T细胞的激活和增殖
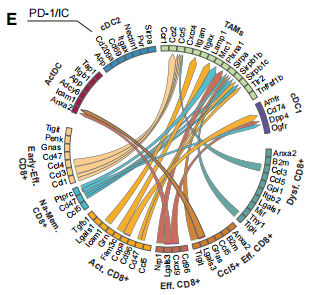
接下来,作者进一步探究BiCE在TME中促进抗肿瘤免疫的模式。通过评估IFN-g和IL-12的细胞内表达水平,作者发现BiCE介导的相互作用细胞获得了生产性促炎DC-T突触的标志性特征,且这两种途径对BiCE的抗肿瘤作用都是单独必需的。为了进一步探索BiCE治疗的分子机制,作者在治疗开始4天后,对来自B16F10荷瘤小鼠的T细胞进行了scRNA-seq分析。通过使用MetaCell算法对数据进行分析,确定了35个元细胞,并确定了9个主要的T细胞亚群。通过对BiCE治疗的小鼠肿瘤及dLN细胞进行配体-受体网络分析,发现BiCE治疗组的DC/T配体-受体相互作用增加。通过分析显示增殖特征的细胞比例,观察到在BiCE处理后,增殖的T细胞显著增加。
 |
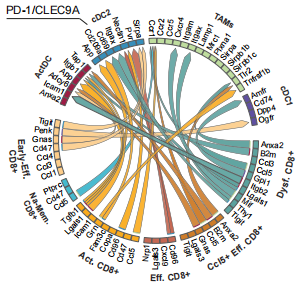 |
这些结果表明,BiCE治疗的效率在早期阶段依赖于IFN-g和IL-12,并调节肿瘤的趋向T-DC相互作用的免疫反应。这种调节导致了TME中CD8+的强大激活、增殖和效应功能。
6. BiCE调节肿瘤浸润淋巴细胞(Tumor-Infiltrating Lymphocyte,TIL)表型
为了更好地理解BiCE导致的强大抗肿瘤疗效的作用模式,作者对肿瘤浸润淋巴细胞(TIL)组成进行了特征分析,并发现BiCE处理的携带B16F10的小鼠肿瘤组织中效应/调节比例显著增加。这表明BiCE治疗介导的DC-T细胞串扰诱导具有独特的促炎分子作用。
为了进一步分析在治疗后期T细胞组成上的区别,作者对处理过的B16F10荷瘤小鼠的T细胞进行了scRNA-seq分析,并得到5个主要的T细胞亚群。对肿瘤T细胞丰度的分析发现BiCE处理后,功能失调的CD8+ T细胞的比例显著下降,treg显著下降,效应CD8+ T细胞亚群增加。因此认为,BiCE治疗将TIL的组成调节成一种更促炎、抗肿瘤的表型,这不同于传统的免疫检查点抑制的效果。
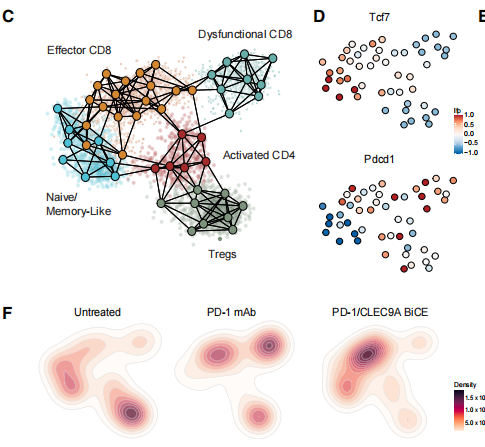
最后,作者探索BiCE治疗导致的TME成分调节的早期事件。通过分析BiCE介导的细胞偶联体中的TCF1水平,以评估TPEX在BiCE治疗开始后的24小时和4天与cDC1的作用。发现在治疗开始4天后,dLN中偶联体TCF1水平在第一天出现的早期富集不再出现,说明BiCE治疗的早期在dLN中主要诱导由TPEX组成的偶联。这可能导致随后在肿瘤中激活的T-DC的增加和CD8+ T细胞增殖的增加。
这些结果突出了BiCE治疗中独特的分子和细胞机制,即在cDC1和TPEX之间产生积极的物理接触,从而增强肿瘤特异性T细胞的增殖和TIL状态和频率的重构,从而有利于肿瘤的消除。
总结
总体而言,这些结果表明,BiCE治疗诱导肿瘤和dLN内相互作用的活化T细胞-DC偶联物,显著改变了肿瘤中效应免疫细胞的数量和细胞状态。在多种侵袭性肿瘤模型中,抗肿瘤免疫的这些细胞和分子变化导致了显著的全身和长期肿瘤控制。因此,本文作者建议BiCE治疗可以作为一种新的免疫治疗方法,通过增强免疫细胞相互作用产生强大的疗效,且对PD-1检查点抑制治疗耐药的肿瘤具有治疗效果。
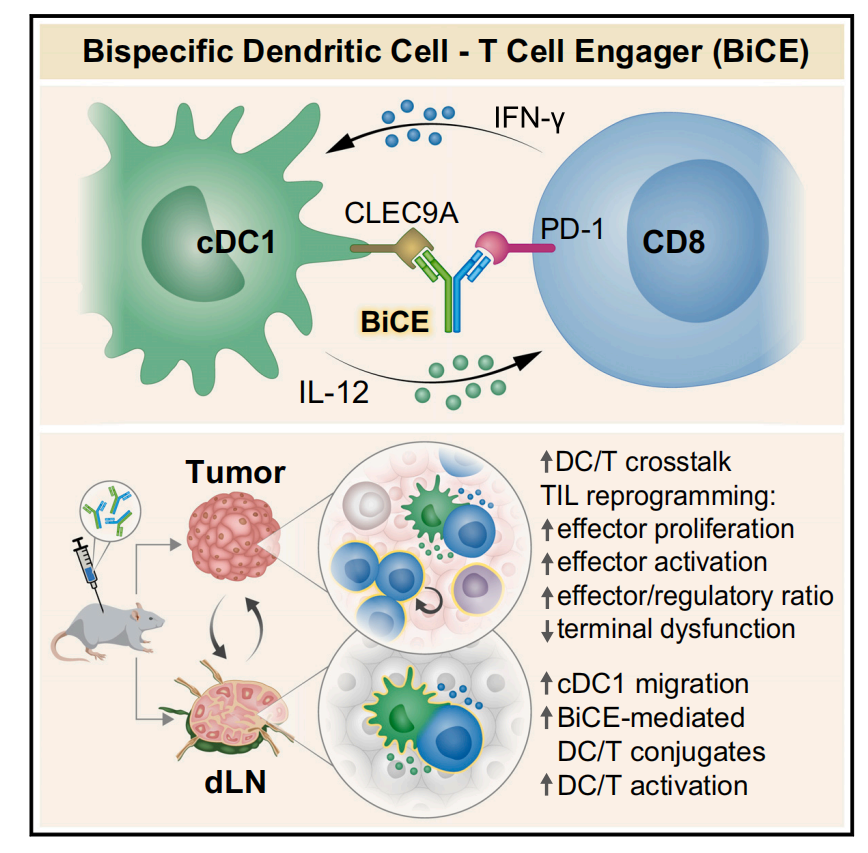
图片总结:一种双特异性抗体促进了传统I型树突状细胞和CD8+ PD-1+ T细胞之间的物理相互作用,增强了apd-1介导的反应所需的免疫通路,并具有强大的抗肿瘤治疗活性。
研究不足
1. 没有直接的实验证明DC是如何被招募,以及如何从dLN转移到肿瘤的;
2. 没有探究BiCE的有效作用时间;
3. 所有的结论都是基于临床前小鼠肿瘤模型,没有临床样本;
4. 有关BiCE机制的几个关键方面仍存在不确定性,例如没有评估CD4+辅助T细胞对BiCE治疗缺陷的贡献,也没有描述CD4+辅助T细胞在BiCE介导的双链中的存在。
参考文献
1. Morad, G., Helmink, B.A., Sharma, P., and Wargo, J.A. (2021). Hallmarks of response, resistance, and toxicity to immune checkpoint blockade. Cell 184, 5309–5337.
2. Sharma, P., Hu-Lieskovan, S., Wargo, J.A., and Ribas, A. (2017). Primary, Adaptive, and Acquired Resistance to Cancer Immunotherapy. Cell 168, 707–723.
3. Garris, C.S., Arlauckas, S.P., Kohler, R.H., Trefny, M.P., Garren, S., Piot, C., Engblom, C., Pfirschke, C., Siwicki, M., Gungabeesoon, J., et al. (2018). Successful Anti-PD-1 Cancer Immunotherapy Requires T Cell Dendritic Cell Crosstalk Involving the Cytokines IFN-g and IL-12. Immunity 49, 1148–1161.e7.
4. Barry, K.C., Hsu, J., Broz, M.L., Cueto, F.J., Binnewies, M., Combes, A.J., Nelson, A.E., Loo, K., Kumar, R., Rosenblum, M.D., et al. (2018). A natural killer–dendritic cell axis defines checkpoint therapy–responsive tumor microenvironments. Nat. Med. 24, 1178–1191.
5. Garris, C.S., Arlauckas, S.P., Kohler, R.H., Trefny, M.P., Garren, S., Piot, C., Engblom, C., Pfirschke, C., Siwicki, M., Gungabeesoon, J., et al. (2018). Successful Anti-PD-1 Cancer Immunotherapy Requires T Cell Dendritic Cell Crosstalk Involving the Cytokines IFN-g and IL-12. Immunity 49, 1148–1161.e7.


 北京市丰台区右安门外西头条10号
北京市丰台区右安门外西头条10号
 010-86738999
010-86738999
 cimr@cimrbj.ac.cn
cimr@cimrbj.ac.cn









 En
En
