
撰文 | 罗斌

2024年11月12日,应首都医学科学创新中心梅林教授邀请,加州大学旧金山分校(University of California, San Fransico)的Roger Nicoll教授在首都医科大学逸夫报告厅带来了一场题为 “A molecular machine for memory”的精彩报告。
Roger Nicoll,神经生物学家,美国国家科学院院士,美国艺术与科学学院院士,现为美国加州大学旧金山分校细胞与分子药理学系荣誉教授。
Roger Nicoll本科在威斯康星州阿普尔顿的劳伦斯大学学习生物学和化学,之后到罗彻斯特大学医学院接受了医学训练,并于1973年获得医学博士学位,在读博期间,他在美国国立卫生研究院学习了一年的电生理学。他于1973年开始在纽约州立大学工作,并在那里与诺贝尔奖获得者 John Eccles 合作。之后他加入了加州大学旧金山分校,并一直留在那里任教。
记忆作为动物最重要的生理功能之一,是动物维持生存繁衍所必需的。记忆在我们的大脑中是如何产生、储存、回溯以及消退的呢?这些重要的问题困扰并吸引着众多的神经科学家们前赴后继、孜孜不倦的研究。
Roger Nicoll以“当我们形成记忆时大脑发生了什么改变?”这一问题引出他的报告,引发听众的思考。中枢神经系统中神经元以电信号网络的传递来快速响应外界的变化,Patricia Goldman-Rakic在1993年通过记录恒河猴额叶皮层神经元活动以及Karel Svoboda在2019年记录小鼠前外侧运动皮层揭示,短时程记忆,如工作记忆,过程中神经元电信号的发放对记忆的形成十分重要。然而,神经元电活动的变化持续时间很短,在时间上很难对于长时程记忆的形成以及记忆的储存发挥作用。因此,除了电生理层面的作用,分子层面对于记忆的形成也应该发挥了十分重要的作用。Roger 引用Donald. O. Hebb 1949年在The organization of Behavior一书中的猜想,认为在记忆的形成过程中,神经元持续性放电会导致突触的增强或减弱,导致分子水平上的改变从而使记忆的形成变得可能。1973年By Bliss 和T. Lomo发表的重要文章首次阐释了在海马离体脑片上发现,短时间的强直刺激可以使突触传递产生长时程增强(Long-Term Potential, LTP)的现象,为后续人们研究记忆的形成提供了重要证据。
Roger表示他的实验室对于LTP的不同阶段(induction, expression, maintenance)都十分感兴趣,并分别对这三个阶段的分子机制进行了深入的研究。

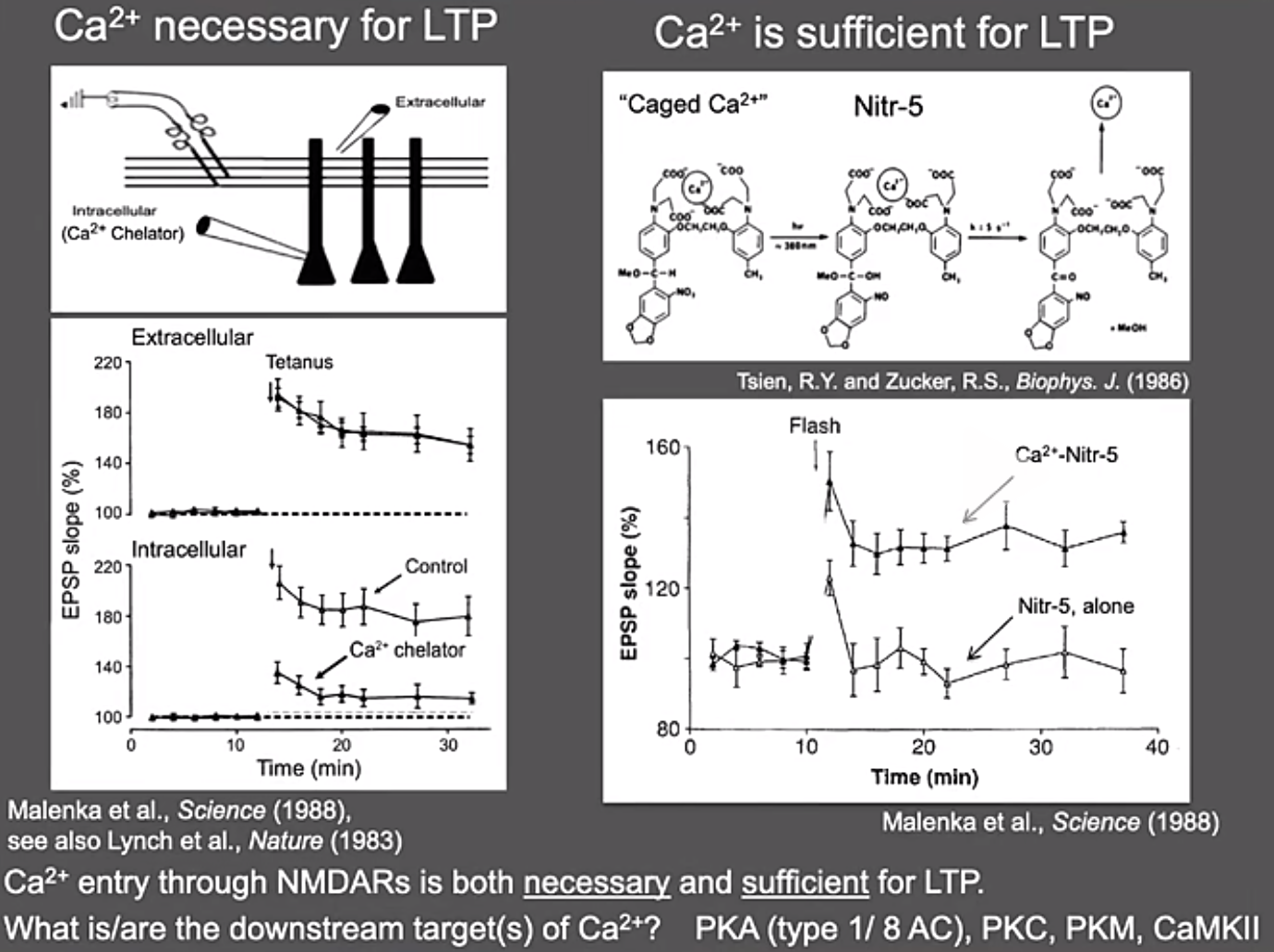
Induction阶段:钙离子作为神经电活动十分重要的离子,其在LTP诱导过程中是否发挥作用呢?Roger首先对钙离子的重要性进行了探究。他们发现在加了钙离子的螯合剂之后,强直刺激引起的单细胞水平上的LTP出现显著的降低,这证明钙离子对于LTP的形成是必要的;为进一步证明钙离子对LTP的形成是充分的,他们利用通常情况下能“锁住”钙离子,光照之后能“释放”钙离子的化学物质Nitr-5处理离体脑片,发现在给予光照之后,提前锁住了钙离子的Nitr-5显著增强了LTP,而没有锁住钙离子的Nitr-5则不行,这证明了钙离子对于LTP的形成是充分的。通过给神经元胞内加入CaMKII的抑制型多肽(inhibitory peptide),他们进一步证明作为钙离子下游的CaMKII对LTP的形成也很重要。反过来,表达持续激活型的CaMKII,同样可以模拟LTP的发生。这一系列实验说明CaMKII是LTP形成的最重要蛋白分子。
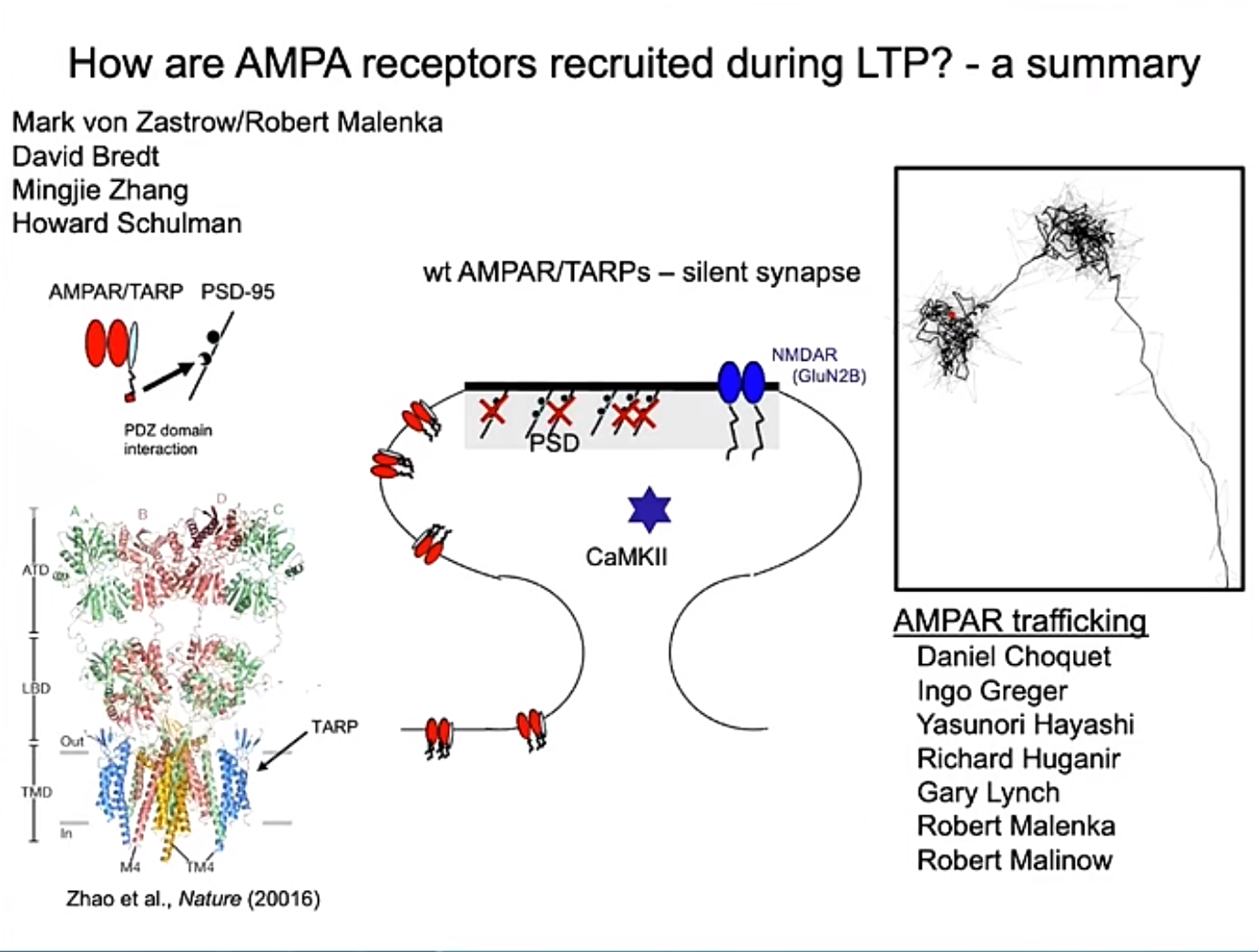
Expression阶段:对于突触前机制与突触后机制在LTP中的作用,领域内有很长一段时间的争议。突触前机制假设认为,LTP的增强是通过突触前神经元释放更多的谷氨酸实现的:LTP后谷氨酸释放概率的提升,导致突触后AMPA受体和NMDA受体的响应都增强,且突触前释放的神经递质无法触发突触后反应在LTP后减少。而突触后机制假设认为,LTP的增强源于突触后AMPA受体的数量或活性增加,而突触前释放不发生显著变化:LTP后AMPA响应增强,而NMDA响应保持不变;原本仅有NMDA受体的突触在LTP后通过增加AMPA受体被“唤醒”。Roger总结了他的实验室关于LTP过程中AMPA受体被招募的机制研究,通过成像、电生理以及分子生化等手段研究,他发现:1. AMPA受体在细胞膜上自由运动,可通过布朗运动进入或离开突触区;2. CaMKII被NMDA受体激活后,与PSD-95蛋白的相互作用解除屏蔽作用,而AMPA受体则通过辅助蛋白TARP与PSD-95结合,被稳定捕获到突触后致密区,这一机制对LTP形成至关重要。
Maintenance阶段:什么样的分子才符合记忆能够被长时间保留的特性呢?Francis Crick 和 John Lisman 提出了记忆分子的最基本要求:首先,必须有一种具有多个相同亚的蛋白质,且这些亚基应具备相互“磷酸化”的能力。其次,该蛋白质的组成亚基可以被新生成的亚基所取代,进行亚基交换,进而发生磷酸化。
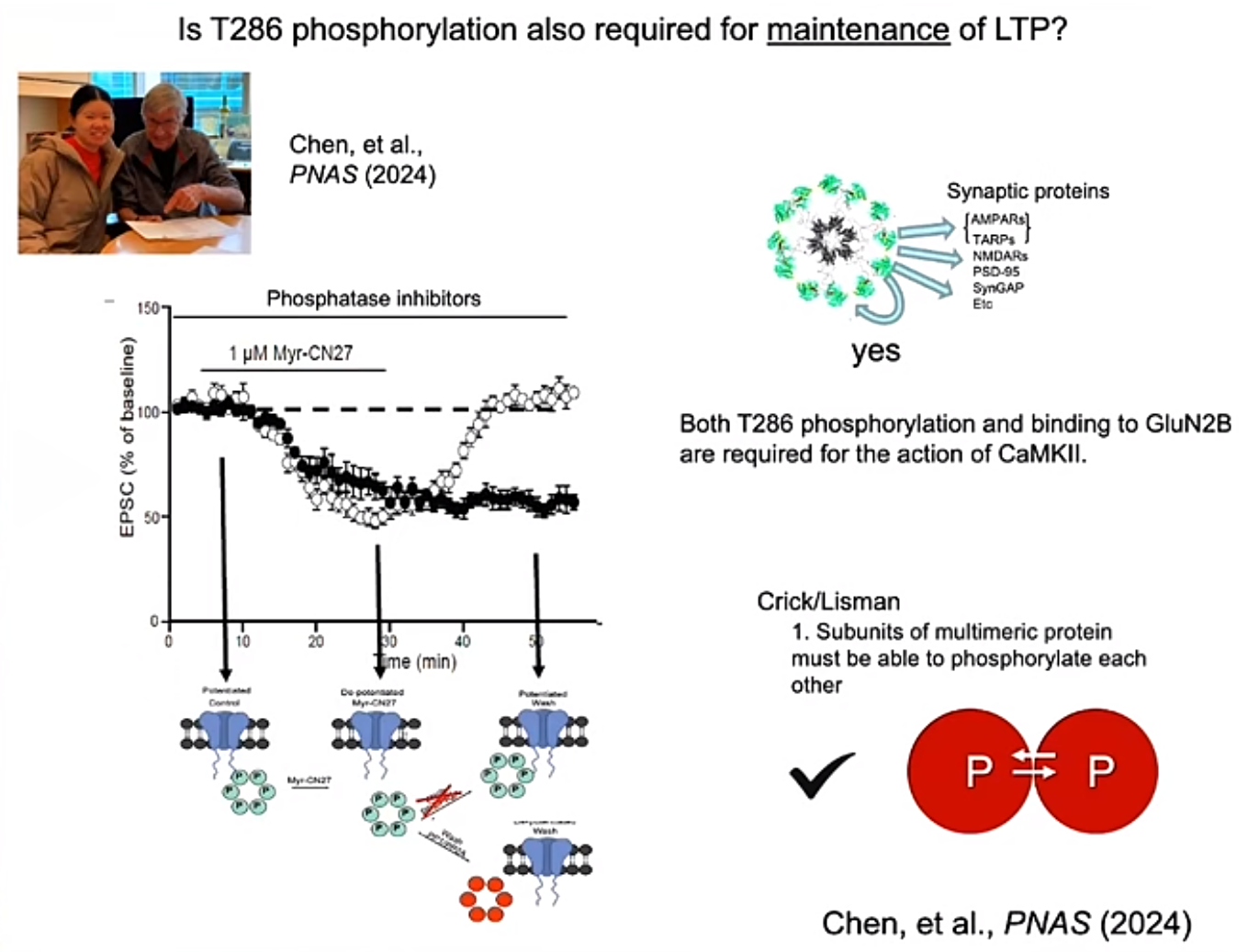
CaMKII被认为在记忆的储存过程中起着至关重要的作用。CaMKII是一个具有12个亚基的多聚蛋白质,在神经元的突触处,它能够通过自磷酸化过程,持续激活,保持其生物学活性。其关键机制之一是钙离子结合钙调蛋白后,促使CaMKII的自磷酸化。但CaMKII的磷酸化是否与LTP有关一直存在争议。Roger实验室使用了一个可以透过细胞膜的棕榈酰化修饰的多肽myr-CN27来抑制CaMKII的磷酸激酶活性,发现CaMKII激酶活性,特别是,286位丝氨酸的磷酸化,以及CaMKII与GluN2B的结合对于LTP maintenance十分重要。
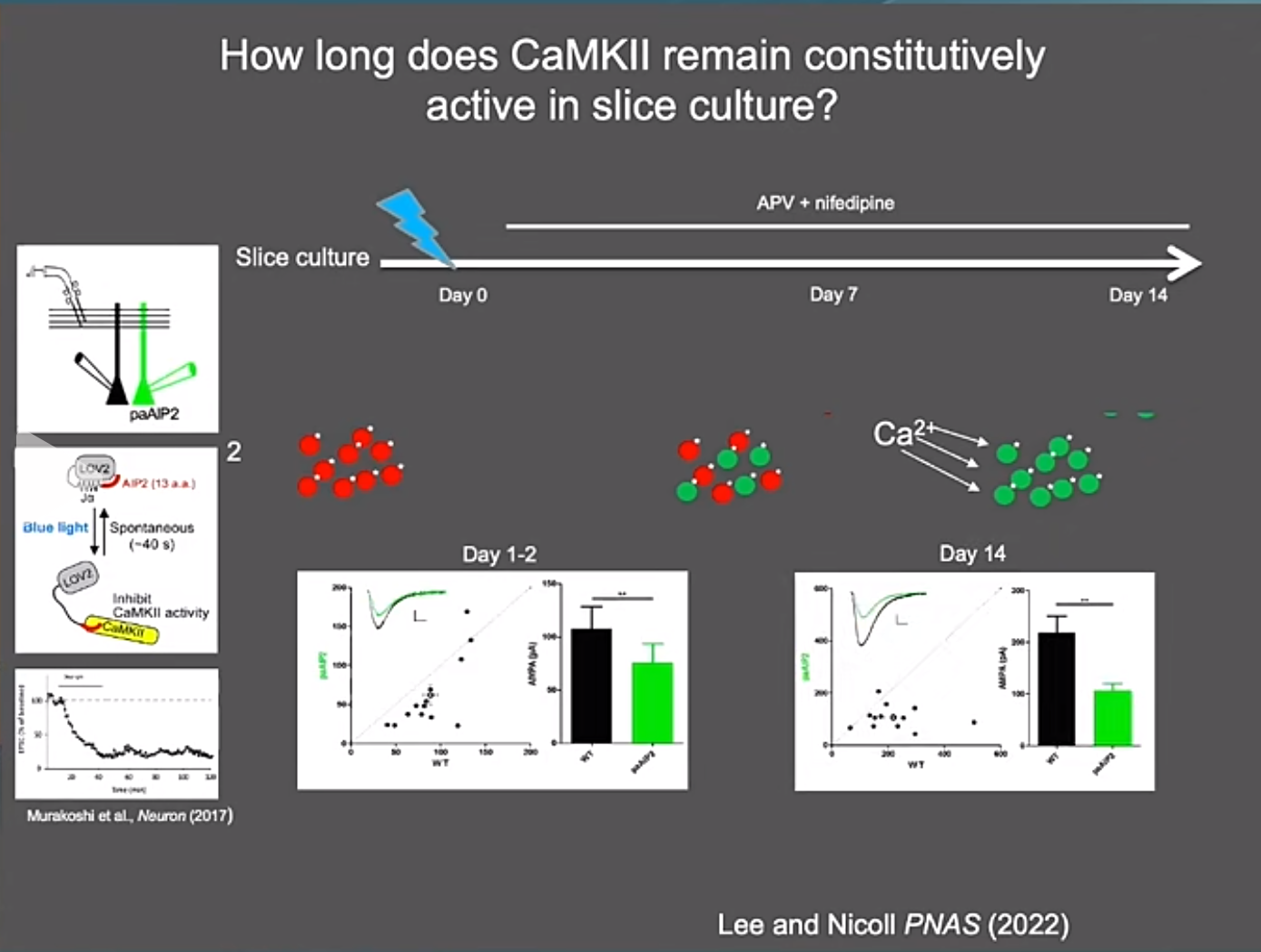
那么CaMKII如何维持LTP呢?虽然CaMKII本身的存在是短暂的,但它的活性却能够通过亚基交换得到延续。2014年一篇发表在eLife上的研究发现CaMKII能够交换其亚基,使得新合成的亚基带有原有的磷酸化状态,这样即便原有的蛋白质已经降解,记忆依然能够维持。这一发现解决了蛋白质寿命与记忆持久性之间的矛盾。Roger实验室利用一种光诱导可逆的CaMKII抑制剂paAIP2,通过给光处理也证明CaMKII持续激酶活性的重要性。并且这一持续的活性即使2周后CaMKII亚基完全更新,也不受影响,且具有之前完整的记忆印记。当然,对于CaMKII磷酸化如何转移到新合成的亚基上,其具体机制还有待进一步研究。
Roger Nicoll教授在本次报告中详细的介绍了LTP的研究发展,向我们展示了不同科学家对于LTP研究的理解思考,并且进一步介绍了他的实验室关于LTP研究的最新进展,使听众对LTP乃至记忆的形成维持分子机制有了更加充分深刻的认识。
参考文献:


 北京市丰台区右安门外西头条10号
北京市丰台区右安门外西头条10号
 010-86738999
010-86738999
 cimr@cimrbj.ac.cn
cimr@cimrbj.ac.cn









 En
En
